Suivi des populations
Suivi des populations
L’approche traditionnelle de la dynamique de population s’articule autour des notions d’effectifs et de taux de croissance. Récemment, les modèles se sont complexifiés et abordent l’ensemble des paramètres démographiques.
Les ornithologues espagnols ont montré que le taux de croissance d'une population de Faucons crécerellettes est influencé par différents facteurs qui sont, pour les principaux, classés par ordre d'importance : le taux de survie adulte, le taux de survie des juvéniles, la productivité moyenne, les fréquences de reproduction des adultes et des subadultes. Les taux d’émigration et d’immigration sont également des paramètres importants mais qui sont plus délicats à évaluer.
En France, les paramètres démographiques de la population (effectif, productivité, survie, fréquence de reproduction…) ont été estimés à partir des données du suivi de la reproduction et d’un programme de baguage.
Le suivi de la reproduction a pour objectif la localisation des sites de reproduction et la détermination de la taille de ponte, du nombre de couples nicheurs, du succès reproducteur et de la productivité. Les principaux sites de colonies sont habituellement occupés d’une année sur l’autre. Cependant, une prospection systématique est nécessaire pour détecter les nouveaux sites colonisés.
Le baguage de la population de la plaine de Crau est mis en œuvre depuis 1994 et depuis 2006, pour la population réintroduite dans l’Aude. La population héraultaise n’est pas baguée du fait de l’impossibilité de capturer les poussins au nid, les cavités étant situées sous les toitures des habitations humaines.
Suivi de la reproduction
L’objectif de ce suivi est de connaître de façon exhaustive les principaux paramètres de reproduction de la population cravenne, soit le nombre de couples nicheurs, la taille des pontes, le taux de réussite, le nombre de jeunes à l’envol, mais aussi de déterminer les principaux problèmes rencontrés par l’espèce en Crau tels que la prédation, les dérangements, la compétition interspécifique…
La taille des pontes
Le contrôle de la taille des pontes est réalisé chaque année. L'objectif est d'obtenir des éléments supplémentaires pour comprendre les variations inter-annuelles du nombre moyen de jeunes à l'envol. Le maximum de précautions est pris pour éviter le dérangement des couveurs. Les nids sont contrôlés une seule fois au cours de la deuxième ou de la troisième semaine d’incubation. Ce contrôle s’effectue au milieu de la journée, en éclairant l'intérieur du nichoir à l'aide d'une lampe torche, sans ouvrir celui-ci : inquiet, l'oiseau s'écarte de sa ponte mais ne quitte pas le nid. Pour la population de la plaine de Crau, la taille moyenne des pontes est de 4.29 œufs par nid. Pour comparaison, Negro (1993) trouve une taille de ponte moyenne égale à 4.11 dans le sud de l’Espagne.
Le sexe-ratio
Un élément important de la dynamique de population est le sexe-ratio. Chez le Faucon crécerellette, il est considéré égal à 1 dans les nichées (Negro & Hiraldo, 1992 ; Tella et al. 1996 ). Pour la plaine de Crau, Mihoub et al. (in prep.) trouvent dans la population subadulte et adulte 0.526 pour les mâles et 0.474 pour les femelles, soit un sexe-ratio proche de l’équilibre (1). La légère différence étant peut-être due à une philopatrie plus élevée chez les mâles que chez les femelles.
La productivité
La productivité est le nombre moyen de jeunes à l’envol par couple nicheur.
En Crau, la productivité varie entre 1.17 et 3.13 selon les années ; la productivité moyenne étant de 1.92. Le facteur principal limitant la productivité est la prédation par les mustélidés et les serpents qui diminue le taux moyen de réussite des couples nicheurs égal à 63 %. Les valeurs annuelles de ce taux de réussite variant entre 43 % et 84 %.
Dans l’Hérault, la productivité varie de 2.36 à 2.78 avec une valeur moyenne de 2.51 (Saulnier, 2013). Cette valeur plus élevée et la plus faible variation constatée sont principalement dues à la meilleure qualité des sites de nidification en milieu urbain qui permet un taux moyen de réussite très élevé égal à 89.90 %. Les valeurs annuelles du taux de réussite varient entre 81.82 % et 95.83 %.
Pour comparaison, Alcazar et al. (2004) estiment la productivité moyenne de la population portugaise à 1.71. Le taux de réussite étant en 2003 de seulement 58 % avec pour les principales causes d’échecs, la prédation des œufs (48 %), la prédation des poussins (9 %), l’abandon des pontes (16 %) et le pillage des nids par l’homme (19 %).

Couleuvre à échelons capturée dans un nichoir

Restes de poussins tués par un putois
Le succès reproducteur
Le succès reproducteur est le nombre moyen de jeunes à l’envol par couple reproducteur réussissant sa reproduction.
Ce succès reproducteur est de 3.03 pour la population cravenne (maximum 3.72 et minimum 2.40) et de 2.80 pour la population héraultaise (maximum 3.00 et minimum 2.65) (Saulnier, 20013 ; Pilard et al., 2013). Cette valeur est légèrement plus élevée pour la population de Crau que pour la population héraultaise. Cependant, la différence est peut-être due à un biais dans la méthode d’observation, puisque l’intérieur des nids est contrôlé en Crau mais pas dans l’Hérault.
La comparaison avec les données des colonies ibériques indique que ce sont des chiffres corrects. En effet, Nunez & Ferrero (2001) trouvent sur une colonie des plaines de Cacérès en Espagne, un succès reproducteur de 2.88 en 1998 et de 3.78 en 1999. Au Portugal, Alcazar et al. (2004) estiment en 2003 le succès reproducteur de la population à 2.91.
Le succès reproducteur rend compte de la quantité et de la qualité des disponibilités alimentaires offertes par les habitats situés autour du site de nidification. Par exemple, Negro et al. (1993) observent sur certaines colonies espagnoles une forte mortalité des poussins (jusqu'à 35 %) qu'ils attribuent à un manque de nourriture. Ce phénomène n'a pas été observé en plaine de Crau où les disponibilités alimentaires semblent élevées.
Rodriguez & Bustamante (2003) mettent en évidence un effet des conditions climatiques sur divers paramètres de la reproduction des colonies en Andalousie. Les précipitations ont en général un effet positif sur les paramètres de la reproduction lorsqu’elles interviennent durant l’hiver précédant la nidification, durant les phases d’installation, de ponte et d’incubation. Par contre, les auteurs constatent un effet négatif des pluies durant la période d’élevage des jeunes. L’effet des pluies de printemps est plus particulièrement positif sur le succès reproducteur car, généralement peu abondantes dans les régions méditerranéennes, elles induisent une croissance forte de la végétation qui favorisent les densités de proies du Faucon crécerellette tels que les orthoptères, insectes principalement herbivores.
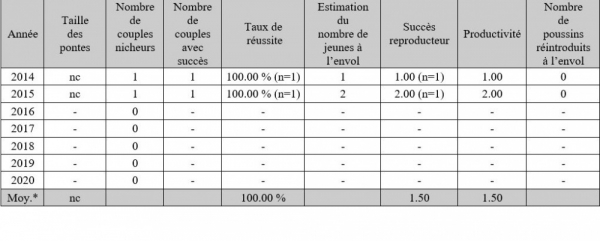
Paramètres reproducteurs du couple gardois en 2014 & 2015
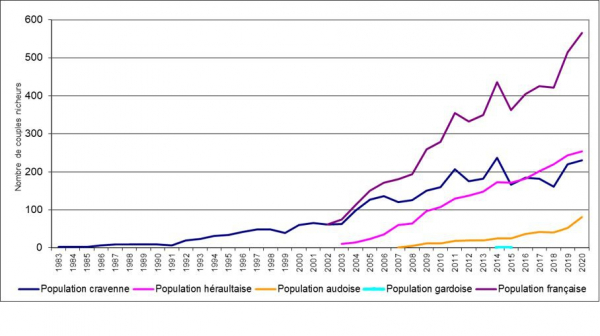
Evolution des effectifs des différentes pouplations françaises depuis 1983
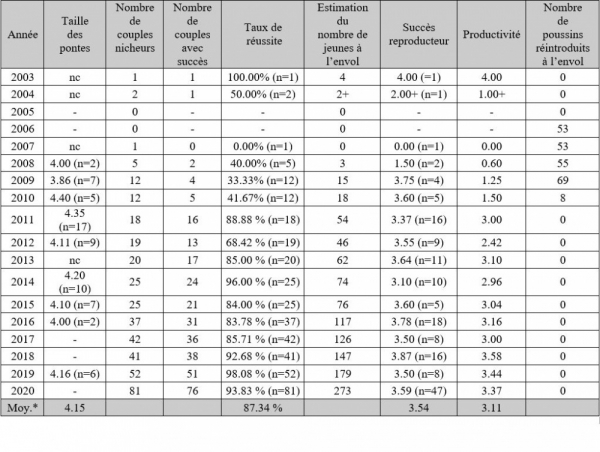
Paramètres reproducteurs de la population de la plaine audoise 2003-2020
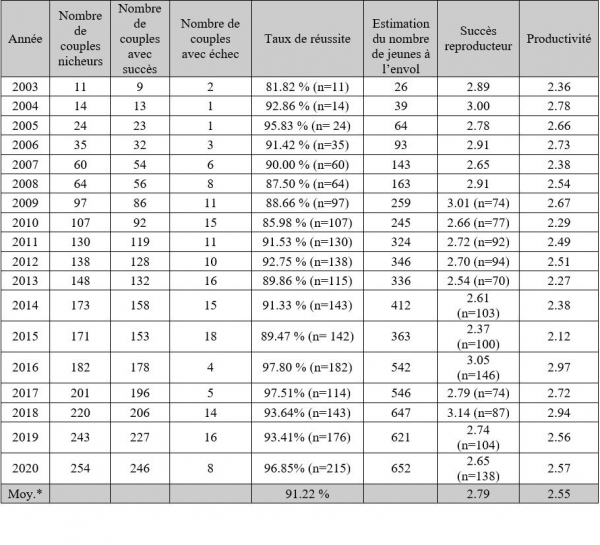
Paramètres reproducteurs de la population centre héraultaise 2003-2020
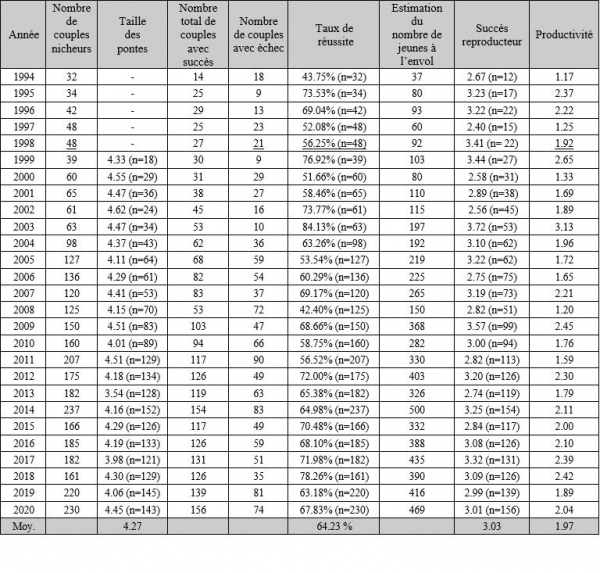
Paramètres reproducteurs de la population de Crau 1994-2020
Programme de baguage

Mesure de l'aile
Le baguage de la population de la plaine de Crau s’effectue depuis 1994. Seuls les poussins sont bagués car les individus adultes ne le sont pas pour éviter un dérangement néfaste à leur reproduction mais aussi, pour différentes raisons qui sont : la difficulté de capture des adultes et le fait que l’espèce acquiert très rapidement l’âge adulte. Les poussins sont capturés de jour ou de nuit dans les cavités de nidification. Ils sont bagués dès que la taille du tarse est suffisamment grande pour la pose d’une bague darvic, soit à partir de l’âge de 20 jours. Sur chaque poussin, nous posons une bague métal et une bague darvic dont le code est lisible à distance grâce à un télescope. Les bagues métal sont fournies par le Centre de Recherche sur la Biologie des Populations d’Oiseaux (CRBPO). Elles ont un diamètre de 7 mm et sont en aluminium. Les bagues en plastique sont de couleur blanche (1994-2007), verte (2008 et 2009) ou jaune (2009-2014) et sont gravées d’un code alphanumérique à 3 caractères lisible à distance à l’aide d’un télescope. Afin d’éviter l’emploi de codes identiques, ces bagues nous sont fournies par la Station biologique de Donana qui développe en Espagne un important programme de baguage sur le Faucon crécerellette. Au moment du baguage, nous déterminons le sexe de chaque poussin et mesurons le poids et la longueur de la 3ème rémige. Ces deux valeurs permettent d’estimer la condition corporelle de chaque poussin.

Bagues plastique et métal
De 1994 à 2020, 4 795 poussins ont été bagués, soit 72.03 % des poussins à l’envol.
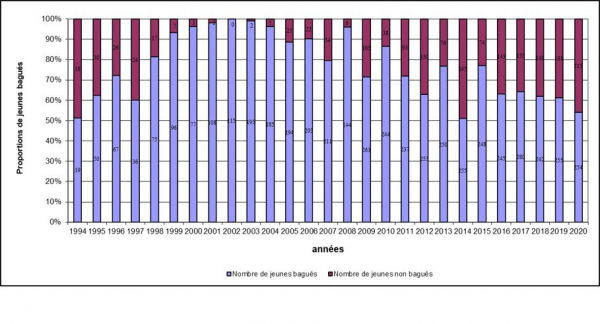
Proportion de jeunes bagués en Crau 1994-2020
La survie des individus
Les taux de survie ont été calculés à partir des programmes MARK et SURGE. Le modèle le plus fiable utilisé est celui où il existe deux classes d’âge (juvénile et adulte), avec une survie juvénile dépendante du temps et une survie adulte constante. La valeur de la survie des adultes est estimée à 0.718 ± 0.013. La valeur de la survie juvénile est estimée à 0.499 ± 0.021 (Mihoub et al., in prep.). La survie adulte est similaire aux valeurs estimées en Espagne: 0.72 ± 0.015 par Serrano et al. (2005) en Aragon et 0.7101 par Hiraldo et al. (1996) en Andalousie. La valeur de la survie juvénile est plus élevée en plaine de Crau que dans le sud de l’Espagne, soit 0.34 estimée par Hiraldo et al. (1996), probablement à cause d’une dispersion juvénile plus importante que pour la population de Crau. De plus, il faut souligner que le caractère constant défini par les modèles pour la survie adulte est relativement fragile.
Mihoub et al. (in prep.) ont montré que la survie juvénile était dépendante de la pluviométrie annuelle en zone sahélienne, pluviométrie qui détermine les densités en Orthoptères, proies principales des Faucons crécerellettes dans ses quartiers d’hivernage ouest africain (Riols, inédit).
On peut s’interroger également de l’impact des pullulations de Criquets pélerins sur les survies juvénile et adulte. En effet, la dernière pullulation constatée au cours des années 2004 à 2006, a coincidé avec des survies particulièrement fortes, pour les juvéniles mais aussi pour les adultes (Pilard & Brun, 2007). Sanchez-Zapata et al. (2007) soulignent l’effet bénéfique de ces pullulations pour l’alimentation des espèces d’oiseaux européens migrateurs trans-sahariens et s’interrogent sur les effets de leur raréfaction sur la dynamique des populations de ces espèces suite au développement de la lutte anti-acridienne (Duranton & Lecoq, 1990).
En Espagne, Serrano et al. (2005) ont montré que la survie adulte augmentait avec la taille des colonies de nidification, soit parallèlement avec la diminution des taux de prédation au nid.
La fréquence de reproduction des individus
En plaine de Crau, le pourcentage des individus subadultes nicheurs (âgés d’1 an) est de 48 % et le pourcentage des adultes nicheurs, dont l’âge est égal ou supérieur à deux ans, est de 88 % (Pilard et al., 2014).
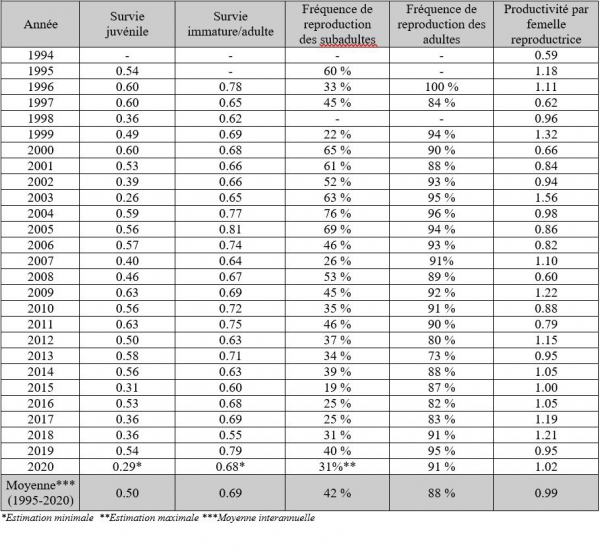
Paramètres de la dynamique de population de Crau 1994-2020
Pour comparaison, Hiraldo et al. (1996) trouvent en Andalousie une fréquence de reproduction adulte similaire (92 %), mais une fréquence de reproduction des subadultes moins élevée (38%), cette dernière étant probablement sous-estimée du fait de la dispersion. Cette différence peut être également due à une meilleure accessibilité des cavités de nidification en plaine de Crau où il existe de nombreux tas de pierres inoccupés situés à proximité immédiate des principales colonies.
Immigration et émigration
La population française est de taille limitée. Elle est relativement isolée et située en limite nord de l'aire de répartition ouest européenne. La plus proche colonie espagnole est la population de Catalogne, forte de 94 couples en 2003 qui est située à environ 100 kilomètres du site de réintroduction audois. Plus éloignée, la population de la vallée de l’Ebre (Aragon) est située à environ 320 kilomètres de la basse plaine de l’Aude et à 470 kilomètres de la plaine de Crau ; elle était forte d’au moins 1 000 couples en 2002 (Pomarol et al., 2004 ; Alcantara de la Fuente, 2004).
Cet état d'isolement géographique relatif est un aspect important de la dynamique de cette population, car il peut théoriquement la fragiliser en limitant les échanges avec les autres populations. En Aragon, Serrano & Tella (2003) ; Serrano et al. (2005) montrent que l’importance des échanges entre sous-populations (une sous-population étant un groupe de colonies) dépend de la distance de séparation et que les échanges se font principalement des plus petites vers les plus grandes.
Le baguage réalisé depuis 1994 en Crau n'apporte que des données partielles sur les échanges existants avec d'autres populations car la proportion d’individus bagués varie selon les populations. Cependant, nous avons observé 2 cas d'émigration de femelles retrouvées nicheuses aux Monégros (Aragon / Espagne) et l'immigration de 5 individus nicheurs porteurs de bagues d'origine étrangère (1 mâle adulte nicheur d’origine inconnue, 2 femelles originaires de Catalogne, 1 mâle adulte originaire de Catalogne, 1 mâle adulte originaire d’Andalousie). A l’exemple d’Hiraldo et al. (1996), une matrice de Leslie a été utilisée pour modéliser la croissance de la population de la plaine de Crau. A partir des paramètres mesurés, le taux de croissance estimé de la population est de 5.3 %. Or, il est très inférieur au taux de croissance moyen observé entre 1993 (24 couples) et 2014 (237 couples) égal à 10.5 %. Cette différence de croissance peut être due à l’arrivée d’oiseaux d’origine extérieure à la population cravenne. Pour l’ensemble de la période 1994 – 2014, la modélisation permet d’estimer ces apports extérieurs à 16 % des individus s’étant reproduit ; par conséquent, 84 % des individus nicheurs seraient originaires de Crau (Pilard et al., 2014).
En région Languedoc-Roussillon, il semble que les cas d’immigration soient plus fréquents. En effet, il existe au moins 9 observations depuis 2002 dont au moins 4 individus sont originaires de Catalogne et 2 de Crau (Rondeau comm. pers. ; Saulnier, 2008).
Cette immigration est bénéfique à double titre, elle améliore le taux de croissance de la population française et la diversité génétique de la population.
Une analyse génétique de différentes populations dont celle de la plaine de Crau (Alcaïde et al., 2008) montre un appauvrissement progressif de la diversité génétique du sud de l’Espagne vers la Catalogne et la France mais celui-ci n’est pas jugé préoccupant pour la conservation de ces deux populations.
En conclusion, il ne semble pas y avoir actuellement de problème majeur d’isolement de la population française même si ce paramètre a pu jouer un rôle plus important par le passé.
Fidélité au site de naissance
La fidélité à la population d’origine, soit la philopatrie, est élevée puisque sur 3 013 poussins bagués en Crau entre 1994 et 2013, 1 013 sont revenus en plaine de Crau, soit 47.2 %. Seulement 8 individus ont été observés nicheurs en dehors de la plaine de Crau : 1 en Aragon, 2 en Catalogne, 3 dans l’Hérault et 2 dans l'Aude. Il est probable que ce chiffre soit sous-estimé du fait de l’absence d’opérations de contrôle dans une grande partie des colonies espagnoles et d'opérations non exhaustives dans l'Hérault.
La dispersion des individus entre la colonie de naissance et la colonie de la première reproduction, soit la dispersion natale, est élevée puisque 81 % (n= 556) des jeunes qui reviennent en Crau s'établissent pour leur première reproduction sur une colonie différente de leur colonie de naissance. A noter qu’en Aragon, Serrano & Tella (2003) trouvent une proportion similaire égale à 83 % et une distance moyenne de dispersion de 7 kilomètres. Par contre, Negro et al. (1997) trouvent en Andalousie une fréquence de dispersion de 43 % et une distance moyenne de 19 kilomètres ; cette dispersion étant identique entre les mâles et les femelles. Par contre, Serrano et al. (2001) trouvent que les femelles (34 %) se dispersent plus que les mâles (19 %) et que cette dispersion diminue avec l’âge des individus.
On note aussi une forte fidélité des oiseaux (81 %) à la colonie de nidification en cas de succès de la reproduction, soit une faible dispersion des reproducteurs, tandis qu'à l'inverse un échec entraîne généralement un changement de colonie. Serrano et al. (2001) démontrent que la probabilité de dispersion des individus nicheurs dépend de deux facteurs principaux, d’une part, du succès de la reproduction et d’autre part, de la proximité des colonies voisines.
Serrano et al. (2004) indique que la dispersion est un facteur important de la dynamique d’une population car elle participe fortement à la sélection ou à l’abandon d’un site de reproduction. Cet auteur constate que le Faucon crécerellette sélectionne prioritairement les colonies de nidification en fonction de l’abondance des individus de la même espèce. Ce paramètre est un indicateur de la bonne qualité du site de nidification plus facile à évaluer par les faucons que la disponibilité alimentaire, la présence de prédateurs ou l’abondance des parasites.
Fidélité au partenaire
Le Faucon crécerellette n’est pas fidèle d’une année à l’autre à son partenaire. Cela est probablement lié à son espérance de vie relativement courte et à sa nidification grégaire. Par contre, afin d’optimiser l’expérience des oiseaux, l’appariement semble s’effectuer en fonction de l’âge. En effet, la sélection du partenaire a lieu au moment de l’installation sur le site de nidification. L’installation des individus âgés s’effectue généralement plus tôt que celle des jeunes individus et par conséquent, les individus d’une même classe d’âge, ayant un même niveau d’expérience, s’apparient généralement entre eux (Pilard, inédit).

Couple de Faucons crécerellettes © Philippe Pilard
Dynamique de la population française
La population française montre depuis 1983 une évolution positive de ses effectifs avec un taux de croissance moyen de 14 % par an. Cependant, la croissance est assez irrégulière dans le temps avec quelques épisodes de fort ralentissement de la croissance et même de diminution des effectifs comme en 1998-1999, ou 2002-2003 et 2007-2008 et des périodes de très forte croissance comme en 1994-1995 et 2004-2006. L’apparente stagnation des effectifs entre 1987 et 1991 est un artefact lié à la présence de colonies non recensées qui n’ont été découvertes en plaine de Crau qu’à partir de 1992. Les variations annuelles des effectifs sont probablement à mettre en relation avec les conditions d’hivernage en Afrique de l’ouest (Mihoub et al., in prep.).
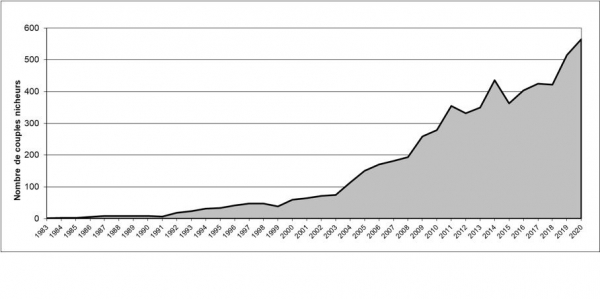
Evolution de la population française depuis 1983
A partir de 2002, la croissance de la population française est renforcée par l’apparition et le développement de la population héraultaise. On constate aussi une croissance plus forte dans l’Hérault qu’en plaine de Crau. En effet, la croissance de la population héraultaise semble soutenue par des apports extérieurs plus importants du fait de sa plus grande proximité avec les populations espagnoles.
De même, à partir de 2007, la croissance est également renforcée par la mise en œuvre d’un projet de réintroduction dans le département de l’Aude.
On constate également, comme on peut s’y attendre pour une espèce coloniale, que l’espèce augmente ses effectifs mais qu’elle augmente peu sa répartition, les individus émigrants préférant apparemment s’installer dans d’autres populations (Monégros, Vallée de l’Ebre, autre population française) que de créer de nouveaux noyaux de colonisation.
Cette évolution positive des effectifs est également constatée dans d’autres pays européens tels que l’Espagne où les effectifs de la population étaient estimés entre 20 000 et 50 000 couples en 1980, entre 4 300 et 5 100 couples en 1990 (Gonzales et al., 1990) et entre 12 000 à 20 000 couples en 2002 (Birdlife International, 2004).